Lakshimi Kiran Chelluri,1 Ravindranath Kancherla,1 Nagaraju Turlapati,2 Sathish Vemuri,1 Tanya Debnath,1 M. Praveen Kumar,2 Syed Sultan Beevi,3 Ratnakar Suguna Kamaraju1
1Transplant Immunology & Stem Cell Lab., Global Hospitals, Hyderabad; 2Zoology Department, Osmania University; 3Centre for Cellular & Molecular Biology, Hyderabad, India
Correspondence: Dr. Lakshmi Kiran Chelluri, Head, Transplant Biology & Stem Cell Lab, Global Hospitals, Lakdi-ka-Pool, Hyderabad, 500 004 (A.P.), India.
Tel. +91.40.30244501 - Fax: +91.40.23244455
E-mail: lkiran@globalhospital.net; apparusu@hotmail.com
Key words: trans-differentiation, islet cells, mesenchymal stem cells, tissue microenvironment.
Acknowledgements: the authors would thank the Department of Science and Technology for the research grant sanction, under Ministry of Science and Technology (DST SR/NM/NAT-03/2006).
Received for publication: 7 February 2011.
Revision received: 2 March 2011.
Accepted for publication: 2 March 2011.
©Copyright L.K. Chelluri et al., 2011
Licensee PAGEPress, Italy
Stem Cell Studies 2011; 1:e5
doi:10.4081/scs.2011.e5
AbstractDirectional transdifferentiation of bone marrow precursor cells assumes beta cell like properties in modified tissue microenvironment. The factors that modify the roles of precursor cells to functional beta cells enabling precise, defined and efficient in vitro differentiation protocols are yet to be conclusive. The study aims at the determination of appropriate induction factors that may aid the robust, reproducible transdifferentiation of rat bone marrow derived mesenchymal stem cells (MSCs) to islet-like cells and enhance their transdifferentiation efficiency. High glucose concentration including nicotinamide, β-mercaptoethanol along with β-cellulin, IGF-1 were able to induce bone marrow precursor cells to islet like clusters ex vivo consistently. The four step induction protocol has enhanced the expression of pancreatic islet cell specific transcription and translational factors detectable by immunocytochemistry viz., pro-insulin, glucagon, somatostatin and polypeptide. The functionality was assessed by the glucose challenge assay followed by animal experiment. The streptozotocin (STZ) induced rats demonstrated significant reduction in glucose levels post islet like cell transplantation (P<0.05). The tropic and the growth factors thus used have a profound impact on the induction of the bone marrow precursors to functional islet like cells. |
β-cell mass is critical in maintaining glucose homeostasis. Deficiency of β-cell and attack on the host cells, due to autoimmunity, impede the insulin secretion.1 Ex vivo generation of β-cell replacement and regeneration of endogenous β-cells are the two major approaches that have been studied extensively during last decade. Various tissue sources and differentiation protocols have been attempted to achieve the insulin levels comparable to mature β-cells.2 Recent researches have reported the generation of insulin by direct differentiation of the precursor cells through over-expression of the transcription factors critical for beta-cell development, such as Pdx-1, Pax-4, Nkx2-2.3,4 Lumelsky5 protocol has been modified by several groups in culture conditions to improve β-cell differentiation. Recapitulation of in vivo signals controlling the tissue microenvironment of the endocrine pancreas is a promising strategy for β-cell generation.6,7 However, the critical point that remains unresolved is the efficient and reproducible differentiation of stem cells into functional insulin secreting β-cells.8,9,10 Hence, an attempt has been made to understand and hypothesize the definitive role of the growth and tropic factors on the tissue microenvironment in culture condition in the development of an efficient and reproducible in vitro differentiation protocol for the generation of β-cells.
Wistar rats weighing about 200 gms and 6 months of age were obtained from National Institute of Nutrition, Hyderabad. All procedures were done in accordance with animal experiment guideline of the Osmania University and Zoology Department clearing Institutional Animals Ethics Committee (IAEC). Cell culture medium L-DMEM (4.5 mmol/L glucose), H-DMEM (25 mmol/L glucose) and fetal bovine serum (FBS) were procured from GIBCO®, Invitrogen Co, Carlsbad, CA, USA. FGF, Trypsin-EDTA, nicotinamide, β-mercaptoethanol, β-cellulin and insulin growth factor (IGF-1) were from Sigma-Aldrich, St. Louis, MO, USA. Anti-Nestin, Pdx-1, Hes-1, Fox A2, Glut-2, Pro-insulin C peptide, somatostatin, glucagon and Pax-6 monoclonal antibodies were from Chemicon, Temecula, CA, USA. LSAB Universal DAB kit was purchased from Dako, Carpinteria, CA, USA.
Bone marrow derived mononuclear cells were isolated from 6 month old male Wistar rat. They were sacrificed by cervical dislocation. The tibias and femurs were recovered by dissection under sterile conditions. The metaphyseal ends of the bones were cut, and the marrow plugs were flushed by passing L-DMEM (GIBCO®) through a needle inserted into one end of the bone. Bone marrow from rats was diluted at 1:1 ratio with normal saline. Mononuclear cells (MNC) fraction was obtained by gradient centrifugation using 1.073 g/mL Ficoll at 1500 rpm for 30 min. The cells were rinsed twice with normal saline and counted using haemocytometer.
The mesenchymal stem cells (MSCs) were cultured as described earlier.11,12 Briefly, the isolated cells were seeded at a density of 1×104/mL in a complete medium containing L-DMEM, FBS (10%) (GIBCO®), penicillin (100 IU/mL), streptomycin (100 IU/mL), gentamycin (50 IU/mL), amphotericin B (2.5 µg/mL) and FGF (10 ng/mL) (Sigma-Aldrich, USA) in polystyrene T25 culture flasks which were not coated with any proteins and were incubated in a humid 5% CO2 incubator at 37°C for 48 hrs. Non adherent cells (hematopoietic cells) were discarded and adherent cells were cultured in complete medium for 10 days, with the medium replaced at 3 day intervals. The cells were trypsinized using Trypsin-EDTA (Sigma-Aldrich, USA) and passaged in T75 flasks until second generation with a confluency of 70-90%. Cultured bone marrow-derived MSCs were then induced for trans-differentiation into functional islet-like cells.
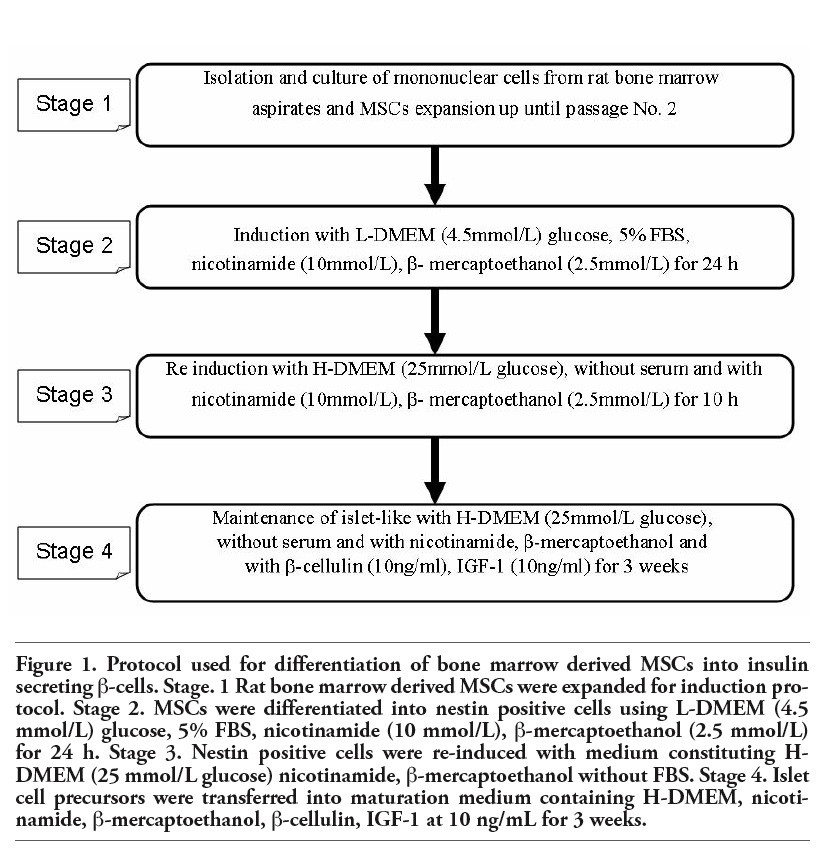
Directed differentiation of MSCs into β-pancreatic cells is achieved through 2-step induction process and single maintenance step. In the first step, MSCs were pre-induced with L-DMEM supplemented with nicotinamide (10 mmol/L), β-mercaptoethanol (2.5 mmol/L) and FBS (5%) (GIBCO®) for 24 h to differentiate into nestin-positive cells. In the second step, nestin-positive cells were re-induced with nicotinamide (10 mmol/L), β-mercaptoethanol (2.5 mmol/L) (Sigma-Aldrich, USA) in serum-free H-DMEM (GIBCO®) for another 10 h to differentiate into premature β-pancreatic islet like cell clusters. In the maintenance step, premature β-pancreatic islet like cell clusters were maintained in long-term culture through addition of IGF-1 (Sigma-Aldrich, USA) and β-cellulin at the concentration of 10 ng/mL in the serum-free culture medium for 3 weeks to obtain mature, insulin secreting islet cells. Cells induced without the addition of nicotinamide, β-mercaptoethanol, IGF-1 (Sigma-Aldrich) and β cellulin was employed as controls (Figure 1).
|
Figure 1. Protocol used for differentiation of bone marrow derived MSCs into insulin secreting β-cells. Stage. 1 Rat bone marrow derived MSCs were expanded for induction protocol. Stage 2. MSCs were differentiated into nestin positive cells using L-DMEM (4.5 mmol/L) glucose, 5% FBS, nicotinamide (10 mmol/L), β-mercaptoethanol (2.5 mmol/L) for 24 h. Stage 3. Nestin positive cells were re-induced with medium constituting H-DMEM (25 mmol/L glucose) nicotinamide, β-mercaptoethanol without FBS. Stage 4. Islet cell precursors were transferred into maturation medium containing H-DMEM, nicotinamide, β-mercaptoethanol, β-cellulin, IGF-1 at 10 ng/mL for 3 weeks. |
Differentiated cells were observed under inverted phase contrast microscope for their morphological changes. Islet cell markers such as Glut-2, Pdx-1, ProC peptides, somatostatin, glucagon (Chemicon) and pancreatic cell differentiation markers such as Fox A2, Hes-1, Pax-6, glucagon (Chemicon) pancreatic polypeptides (PPP) were detected with immunocytochemical techniques using appropriate antibodies. Differentiated cells adherent to poly-L-Lysine coated slides were fixed with cold methanol, washed thrice with PBS and incubated with islets cell markers and pancreatic cell differentiation markers as per the manufacturer’s instructions, after permeation with 0.1% Triton X100 and blocking with 4% BSA. Briefly, they were incubated with primary antibodies against pancreatic cell developmental markers such as, anti-FoxA2, anti-Glucagon, anti-Hes-1, anti-Pax-6, anti-PPP (1:1000 dilution) and pancreatic islet cell developmental markers such as, anti-Glut-2 (1:500), anti-Pdx-1 (1:200), anti-glucagon (1:500), anti-Somatostatin (1:500), anti-proinsulin C-peptide (1:200) at 4°C overnight. They were further treated with secondary antibodies, which were then followed with the chromogen substrate reaction. All the slides were counter stained with hematoxylin. Immunoreactive cells were visualized for their positive/negative staining using inverted phase contrast microscope (magnification x400). The normal islet cells from the rat were used as positive controls (after ensuring in H&E stain) and undifferentiated MSCs were used as negative controls.
The amount of immunoreactive insulin secreted from differentiated and undifferentiated cells after 48 hrs of induction was determined by ultrasensitive one-step immunoenzymatic (sandwich) insulin assay as per the manufacturer's instructions. Briefly, Insulin levels in culture supernatant were determined using mouse monoclonal anti-insulin alkaline phosphatase conjugate and paramagnetic particles coated with mouse monoclonal anti-insulin antibody. A chemiluminescent substrate, Lumi-Phos* 530 was added to the reaction tube and the light generated by the reaction was measured with a luminometer. The photon production is proportional to the amount of conjugate bound to the solid support. The amount of analyte in the supernatant was determined by means of a stored, multi-point calibration curve.
Diabetic animal models were made according to the standard procedure with modifications. In all, 12 Wistar rats (weighing about 200 grams) were intravenously injected with 50 mg/L streptozotocin (STZ) through caudal veins, and glucose levels were tested a week later with Roche ACCU-CHEK glucose tests (Mannheim, Germany). Of the 12 animals, one did not develop diabetes, one died and two reverted back, thus reducing the sample size to 5 animals in the treatment group and 3 in the untreated group.
The glucose controlling functionality of MSCs differentiated islet-like cells were evaluated using 5 diabetic rats (treatment group) along with three sham (untreated group), as compared to undifferentiated cells. After ascertaining the previous reported data, a consensus dose and the route was adopted to perform the above mentioned experiment. Differentiated islet-like cells were trypinized and viability was evaluated by trypan blue exclusion assay before injection into experimental animals. Cell viability was found to be 98% in all the samples processed. Each rat was administered in peritoneum 1×106 cells/mL of differentiated cells, in 0.5 mL of saline as medium. The sham group received only saline as their treatment. Glucose level was measured about two weeks after the therapy.
The results were expressed as mean ± standard deviation (SD). The statistical significance of differences was assessed by Student’s t-test. In all comparisons, a value of P<0.05 was considered statistically significant. Correlation coefficient was used to establish the relationship between the extents of differentiation with reference to the insulin release. Percentage efficiency of differentiation was calculated as number of differentiated cells/number of MSCs per field × 100.
MSCs were spindle shaped and fibrocyte-like on day 14 with 90% confluency. The MSCs characterization and differentiation potential has already been established as previously published.11,12 On differentiation, MSCs transformed into nestin-positive cells and on further induction, they differentiated into typical grape like clusters. Immunocytochemistry was performed to profile these islets-like cells for endocrine differentiation mediated regulatory markers. Nestin was regarded as an important pre-induction phenotypic marker for islet cell differentiation, and its expression was tested. The experimental approach of inducing precursor cells with H-DMEM (GIBCO®) medium along with Nicotinamide, β-mercaptoethanol, without FBS (GIBCO®) has been reproduced several times. These differentiated islets like cells were also maintained in long term culture with modifications in the culture conditions by the addition of insulin growth factor-1 (IGF-1) (Sigma-Aldrich) and β cellulin (Figure 2).
The cells were further processed using islet cell marker and pancreatic cell markers for endocrine releasing hormones, translational and transcriptional factors. The cells were stained positive for endocrine markers (specifically β cell markers) like Pdx-1, Pro-insulin C, and Glut-2 (Chemicon), stained negative for the alpha cell markers like glucagon and delta cell markers like somatostatin respectively (Figure 3). The early pancreatic differentiation markers such as Hes-1 was negative, while Fox A2 (Chemicon), an earlier differentiation marker demonstrated partial positivity. The undifferentiated cells were negative for both of these markers. The late differentiation markers such as Glut-2 (Chemicon), insulin were positive suggesting the transdifferentiation of MSCs to β islet like cells (Figure 4).
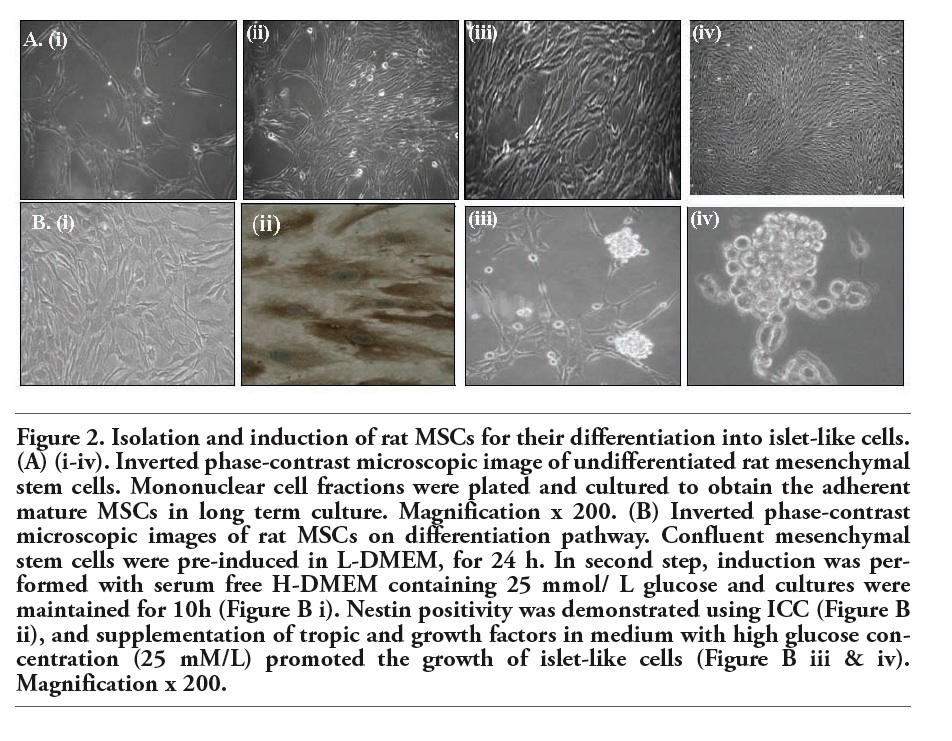 |
Figure 2. Isolation and induction of rat MSCs for their differentiation into islet-like cells. (A) (i-iv). Inverted phase-contrast microscopic image of undifferentiated rat mesenchymal stem cells. Mononuclear cell fractions were plated and cultured to obtain the adherent mature MSCs in long term culture. Magnification x 200. (B) Inverted phase-contrast microscopic images of rat MSCs on differentiation pathway. Confluent mesenchymal stem cells were pre-induced in L-DMEM, for 24 h. In second step, induction was performed with serum free H-DMEM containing 25 mmol/ L glucose and cultures were maintained for 10h (Figure B i). Nestin positivity was demonstrated using ICC (Figure B ii), and supplementation of tropic and growth factors in medium with high glucose concentration (25 mM/L) promoted the growth of islet-like cells (Figure B iii & iv). Magnification x 200. |
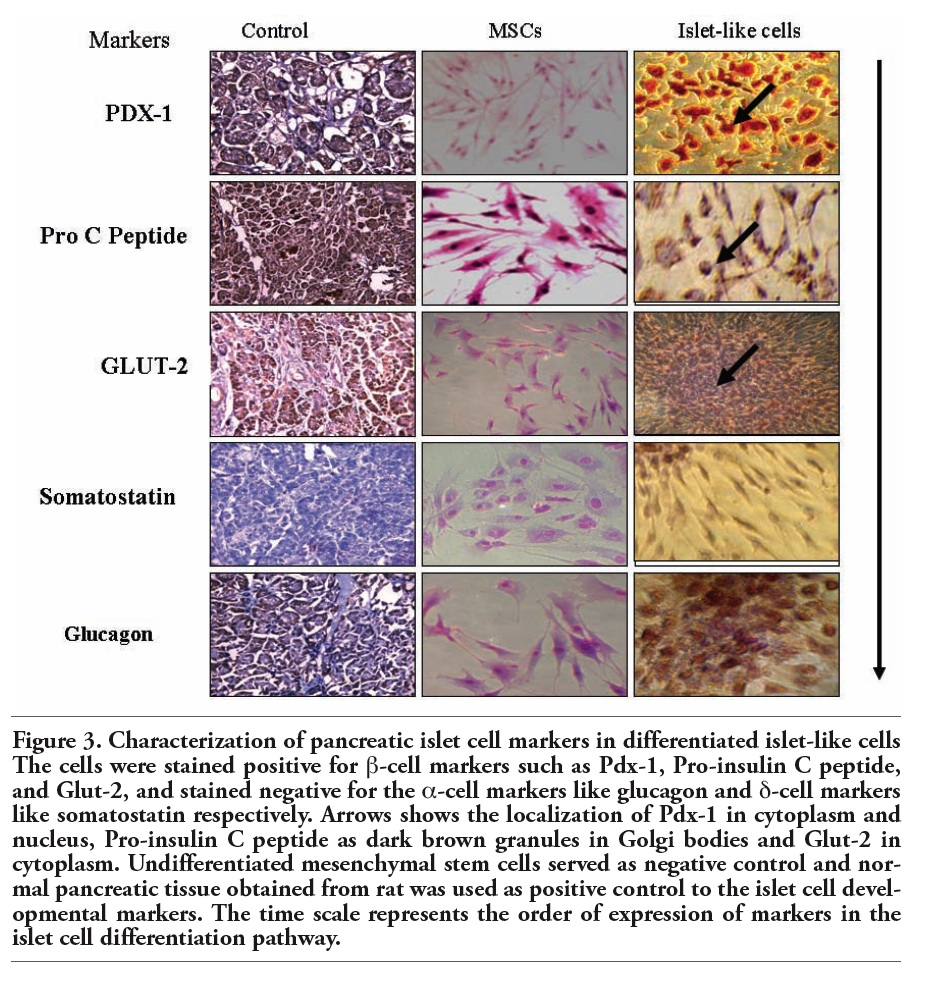 |
Figure 3. Characterization of pancreatic islet cell markers in differentiated islet-like cells The cells were stained positive for β-cell markers such as Pdx-1, Pro-insulin C peptide, and Glut-2, and stained negative for the α-cell markers like glucagon and δ-cell markers like somatostatin respectively. Arrows shows the localization of Pdx-1 in cytoplasm and nucleus, Pro-insulin C peptide as dark brown granules in Golgi bodies and Glut-2 in cytoplasm. Undifferentiated mesenchymal stem cells served as negative control and normal pancreatic tissue obtained from rat was used as positive control to the islet cell developmental markers. The time scale represents the order of expression of markers in the islet cell differentiation pathway. |
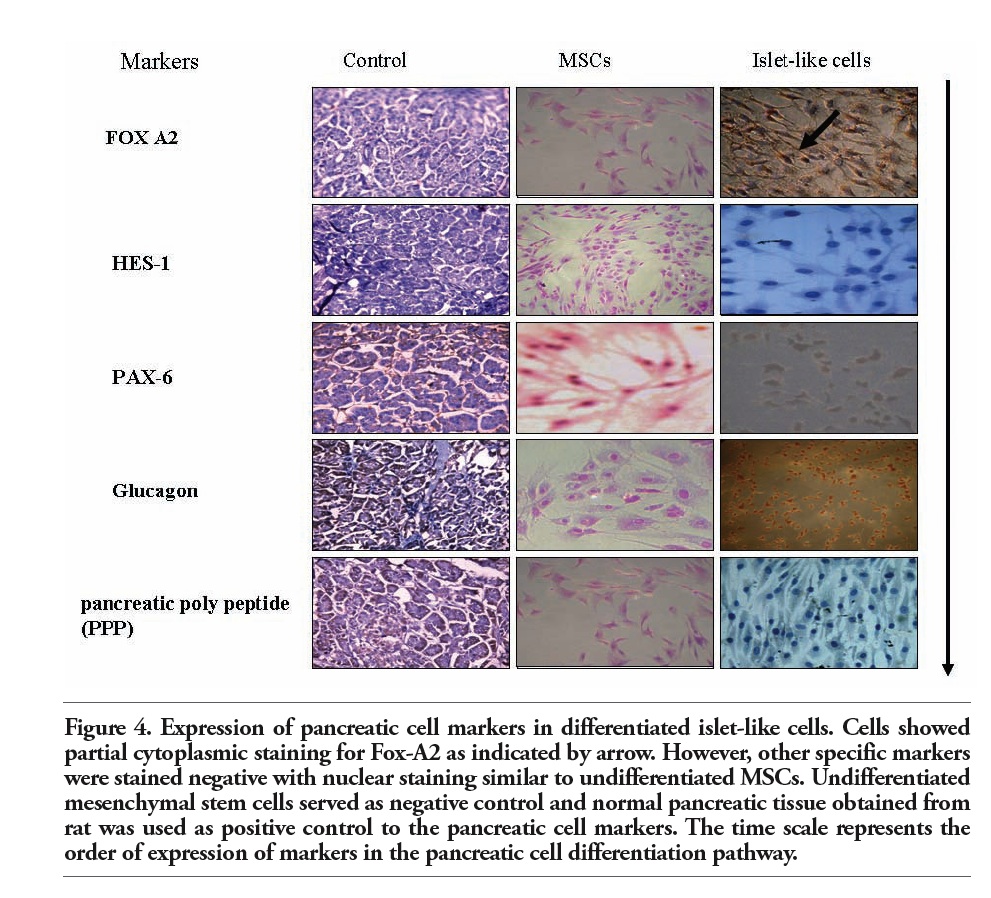 |
Figure 4. Expression of pancreatic cell markers in differentiated islet-like cells. Cells showed partial cytoplasmic staining for Fox-A2 as indicated by arrow. However, other specific markers were stained negative with nuclear staining similar to undifferentiated MSCs. Undifferentiated mesenchymal stem cells served as negative control and normal pancreatic tissue obtained from rat was used as positive control to the pancreatic cell markers. The time scale represents the order of expression of markers in the pancreatic cell differentiation pathway. |
In order to evaluate the functionality of the differentiated islet cells, the cultures at passage 2 stage and 7 day post induction (on microscopic observation for viable and consistent islet like cells) were subjected to various concentrations of glucose in the media and the corresponding differentiated cells and the insulin levels were correlated. Undifferentiated MSCs were also subjected to varied concentrations of glucose and corresponding insulin release was estimated. The release of insulin was significant at P<0.05 at high glucose concentrations from differentiated cells as compared to MSCs alone (Figure 5).
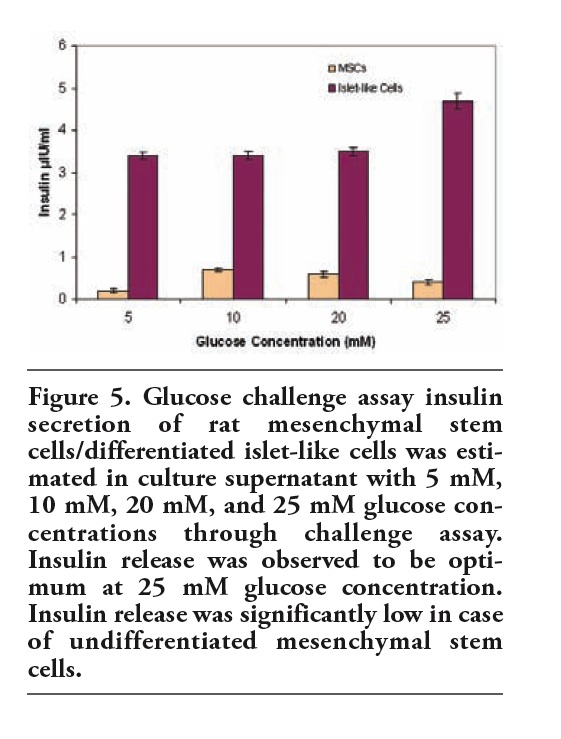 |
Figure 5. Glucose challenge assay insulin secretion of rat mesenchymal stem cells/differentiated islet-like cells was estimated in culture supernatant with 5 mM, 10 mM, 20 mM, and 25 mM glucose concentrations through challenge assay. Insulin release was observed to be optimum at 25 mM glucose concentration. Insulin release was significantly low in case of undifferentiated mesenchymal stem cells. |
To further determine and establish a correlation between efficiency of insulin release in comparison with number of differentiated islets, correlation coefficient was calculated. Different fields were selected in the culture plates and differentiated islet like cells were scored. Insulin release and total number of differentiated islet were considered to computate correlation coefficient. The measure of interdependence was 0.8; demonstrating that corresponding variables closely vary together in the same direction suggesting a positive correlation of the insulin release with reference to the differentiated islets-like cells (Table 1).
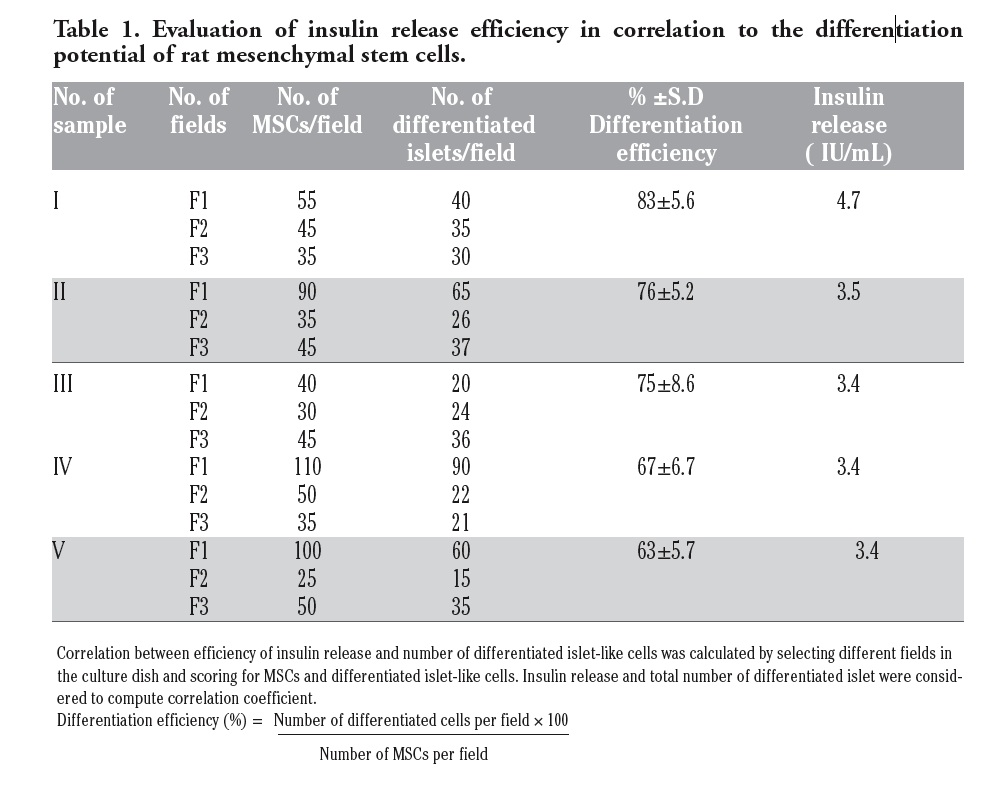 |
Table 1. Evaluation of insulin release efficiency in correlation to the differentiation potential of rat mesenchymal stem cells. |
Glucose levels of 8 rats about 14 days after the injection of differentiated cells/saline are shown in Table 2. Glucose level was found to be reduced significantly in the treated group when compared with their glucose level before the treatment. Based on statistical significance it could be suggested that differentiated islet-like cells has a positive effect on the diabetic status of the STZ induced rat.
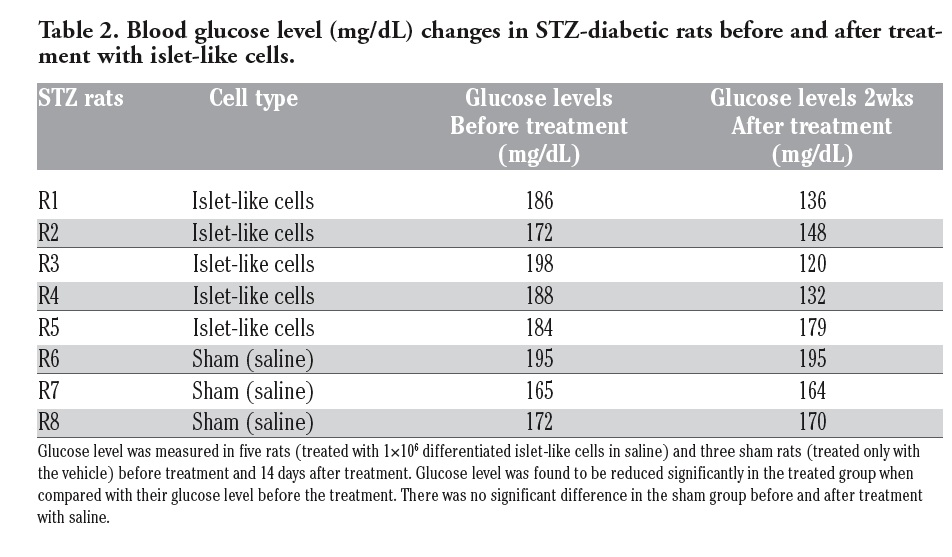 |
Table 2. Blood glucose level (mg/dL) changes in STZ-diabetic rats before and after treatment with islet-like cells. |
The current protocols for efficient transdifferentiation are not yet optimized for a number of reasons viz., pleiotropic effects induced by morphogens, the complexity of the signaling pathways involved, and the likelihood of cross talk between pathways including sources of renewable β-cells.13 Several studies have demonstrated variable degree of successful islet cell differentiation from sources including pancreatic ductal cells to exogenous expression of β-cell transcription factors including several others.14,15 There is ample data support in animal model experimentation that the bone marrow derived precursor stem cells can themselves mediate by circulating signals to the neuro-endocrine compartment of the pancreas and correct the β-cell deficiency in the presence of host vascular endothelial growth factor (VEGF). Further, any substitute for primary islets of Langerhans will require some minimum essential properties if it is to be used for transplantation therapy of diabetes. Vast numbers of replacement of β-cells to make significant therapeutic impact could be achieved using adult tissue such as bone marrow.16,17 The rationale of the adult bone marrow as the tissue source for islet cell differentiation is to circumvent the paucity, availability of functional islets for the transplantation purpose. MSCs are easily expandable to the desired dose and can be grown efficiently in an ex vivo tissue microenvironment.18,19 Furthermore, establishment of efficient and reproducible protocols will be facilitated by these expandable cell populations in order to study on the intracellular signaling pathways that regulate and direct the developmental transitions using appropriate tropic and growth factors. As proposed in Figure 1, the current protocol of using nicotinamide and β mercaptoethanol (Sigma-Aldrich) as a pre-induction step has promoted the endocrine differentiation in the bone marrow derived precursor cells and has repeatedly demonstrated as a maturation factor. Nicotinamide (Sigma-Aldrich) as a morphogen may demonstrate some of its effects mediated through the inhibition of poly (ADP-ribose) synthase. This may cause chromatin rearrangements and changes in gene transcription.20,21,22 Nicotinamide and β mercaptoethanol (Sigma-Aldrich), both as notch repressors along with the high glucose concentrations have the potential to adjust the adult precursor cells to the islet cell tissue microenvironment. Morphologically, the precursor cells in the pre-induction medium demonstrates the chromatin rearrangements and show positivity to nestin marker.23 The nestin positivity thus observed by us confers with other reports with particular reference to preliminary induction phase bearing similarity in the generation of neurons and islets by using gene adjusting pathways.24 Ex vivo differentiation of bone marrow precursor cells to insulin producing β-cells is a complex process with a cascade of gene activation events controlled by the transcription and translational factors including basic helix-loop-helix (bHLH) proteins.25 The ability to identify the lineage markers will allow us to determine beta cell translational factors in the differentiation cascade and fortify the islet cell differentiation protocol.26,27 Pdx-1 (Chemicon), expressed in nucleus is critical in the transactivation of insulin gene. The β-cell-specific glucose transporter, Glut-2 (Chemicon) is essential in maintaining β-cell homeostasis. Approximately 90% of β-cells and 15% of δ-cells in adult islets express Pdx-1.28 Hence, the expression of these markers can be used as key indicator of β-cell functionality. The dual activation of Pdx-1, as a pancreas commitment factor during embryogenesis and as a regulator of islet cell physiology as observed in the results of the Immunocytochemistry (ICC), underscores the unique role of Pdx-1 in maintaining the function of human pancreatic endocrine cells.29 Pro-insulin consists of three domains: an amino-terminal B chain (30 amino acids), a carboxy-terminal A chain (21 amino acids) and a connecting peptide in the middle known as the C-Peptide. Mature form of insulin is released within the endoplasmic reticulum, as a result of exposure of the pro-insulin to several specific endopeptidases and the excision of C-peptide. Insulin and free C-peptide are packaged in the Golgi into secretory granules, which accumulate in the cytoplasm. In the pancreatic β-cells, C-peptide and mature insulin are present in equimolar ratios and co-localize in the secretory granules as demonstrated by the dark brown granules by ICC.30 Pro-insulin C peptide thus localized in the Golgi body is critical for the insulin release as evidenced by the glucose challenge assay and animal experiment. The differentiation protocol thus has been able to obtain functional islets that could be used for transplantation purpose. The pancreatic islet markers have more or less demonstrated negative staining except for Fox A2 (Chemicon), which was present partially in the cytoplasm. This could be attributed to some cells demonstrating the pancreatic differentiation potential and may not be of major concern.
The gold standard for defining β-cell function is glucose responsiveness. The functionality of the differentiated islets when challenged with varied concentrations of glucose levels in the media has resulted in appropriate release of insulin levels.31,32,33,34 It was observed that the rat MSCs showed 60-70% differentiation potential from the primary precursor cells. The required number of differentiated islets for transplant purpose will majorly be dependent on the initial seeding of MSC population.
MSCs alone on glucose challenge assay has demonstrated a dynamic state of the cells with conditional release of the insulin as shown in Figure 5, thereby suggesting the possible role of bone marrow derived MSCs as a combination mode of therapeutic modality in conjunction with differentiated islet cells.35,36 Insulin secreted from the differentiated islet-like cells ameliorated streptozotocin (STZ)-induced diabetes in experimental animals. The replaced cells having the ability to synthesize, store and release insulin as and when required to the dynamic changes of ambient glucose challenge under ex vivo and in vivo condition has been achieved with the optimization of the induction factors. In conclusion, the modified protocol successfully demonstrates a potential β-cell formation in a controlled environment. It is therefore critical to focus on generating populations of cells that possess the phenotype of β-cells with greater immunocompetency to survive and sustain for a prolonged duration of time.
1. Oliver-Krasinski JM, Stoffers DA. On the origin of the beta cell. Genes Dev 2008;22:1998-2021.[PubMed]
2. Ishiyama N, Kanzaki M, Seno M, et al. Studies on the betacellulin receptor in pancreatic AR42J cells. Diabetologia 1998; 41:623-8.[PubMed]
3. Bonner-Weir S, Weir G. New sources of pancreatic beta cells. Nat Biotechnol 2005;23:857-861.[PubMed]
4. Shiroi A, Ueda S, Ouji Y, et al. Differentiation of embryonic stem cells into insulin producing cells promoted by Nkx2-2 gene transfer. World J Gastroenterol 2005;11:4161-6.[PubMed]
5. Lumelsky N, Blondel O, Laeng P, et al. Differentiation of embryonic stem cells to insulin-secreting structures similar to pancreatic islets. Science 2001;292:1389-94.[PubMed]
6. Sosa-Pineda B, Chowdhury K, Torres M, et al. The Pax4 gene is essential for differentiation of insulin-producing beta cells in the mammalian pancreas. Nature 1997386:399-402.[PubMed]
7. Murtaugh LC, Melton DA. Genes, signals, and lineages in pancreas development. Annu Rev Cell Dev Biol 2003;19:71-89.[PubMed]
8. Edlund H. Developmental biology of the pancreas. Diabetes 2001;50:S5-9.[PubMed]
9. Bonner-Weir S. Islet growth and development in the adult. J Mol Endocrinol 2000;24:297-302.[PubMed]
10. Massagué J, Chen YG. Controlling TGF-beta signaling. Genes Dev 2000;14:627-44.[PubMed]
11. Koppula PR, Chelluri LK, Polisetti N, Vemuganti GK. Histocompatibility testing of cultivated human bone marrow stromal cells - a promising step towards pre-clinical screening for allogeneic stem cell therapy. Cell Immunol 2009;259:61-5.[PubMed]
12. Subbaiah GPV, Adavi V, Chelluri LK, et al. Preliminary report on the safety, efficay and functional recovery of spinal cord injury with autologous bone marrow derived mesenchymal stem cells - a clinical trial. Internet J Spine Surg 2009;5:Num. 1[Full-Text]
13. Nikolova G, Jabs N, Konstantinova I, et al. The vascular basement membrane: a niche for insulin gene expression and Beta cell proliferation. Dev Cell 2006;10: 397-405.[PubMed]
14. Brons IG, Smithers LE, Trotter MW, et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature 2007;448:191-5.[PubMed]
15. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006;126:663-76.[PubMed]
16. Burns CJ, Persaud SJ, Jones MP. Stem cell therapy for diabetes: do we need to make beta cells? Jour Endocrinol 2004;183;437-43.[PubMed]
17. Lu Y, Wang Z, Zhu M. Human bone marrow mesenchymal stem cells transfected with human insulin genes can secrete insulin stably. Ann Clin Lab Sci 2006;36: 127-36.[PubMed]
18. Hao E, Tyrberg B, Itkin-Ansari P, et al. Beta-cell differentiation from nonendocrine epithelial cells of the adult human pancreas. Nat Med 2006;12:310-6.[PubMed]
19. Guz Y, Nasir I, Teitelman G. Regeneration of pancreatic beta cells from intra-islet precursor cells in an experimental model of diabetes. Endocrinology 2001;142:4956-68.[PubMed]
20. Vaca P, Berná G, Martin F, Soria B. Nicotinamide induces both proliferation and differentiation of embryonic stem cells into insulin producing cells. Transplant Proc 2003;35:2021-3.[PubMed]
21. Otonkoski T, Beattie GM, Mally MI, et al. Nicotinamide is a potent inducer of endocrine differentiation in cultured human fetal pancreatic cells. J Clin Invest 1993;92:1459-66.[PubMed]
22. Jensen J, Heller RS, Funder-Nielsen T, et al. Independent development of pancreatic alpha- and beta-cells from neurogenin3-expressing precursors: a role for the notch pathway in repression of premature differentiation. Diabetes 2000;49:163-76.[PubMed]
23. Hart A, Papadopoulou S, Edlund H. Fgf10 maintains notch activation, stimulates proliferation, and blocks differentiation of pancreatic progenitor cells. Dev Dyn 2003;228:185-93.[PubMed]
24. Zulewski H, Abraham EJ, Gerlach MJ, et al. Multipotential nestin-positive stem cells isolated from adult pancreatic islets differentiate ex vivo into pancreatic endocrine, exocrine, and hepatic phenotypes. Diabetes 2001;50:521-33.[PubMed]
25. Lee JE, Hollenberg SM, Snider L, et al. Conversion of Xenopus ectoderm into neurons by NeuroD, a basic helix-loop-helix protein. Science 1995;268:836-44.[PubMed]
26. Shapiro AMJ, Lakey JR, Ryan EA, et al. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N Engl J Med 2000;343:230-8.[PubMed]
27. Eiselein L, Schwartz HJ, Rutledge JC. The challenge of type 1 diabetes mellitus. ILAR J 2004;45:231-6.[PubMed]
28. Kim SK, Hebrok M. Intercellular signals regulating pancreas development and function. Genes Dev 2001;15:111-27.[PubMed]
29. Cho YM, Lim JM, Yoo DH, et al. Betacellulin and nicotinamide sustain PDX1 expression and induce pancreatic beta-cell differentiation in human embryonic stem cells. Biochem Biophys Commun 2008;366:129-34.[PubMed]
30. Molinete M, Irminger JC, Tooze SA, Halban PA. Trafficking/ sorting and granule biogenesis in the beta-cell. Semin Cell Dev Biol 2000;11:243-51.[PubMed]
31. Shiroi A, Yoshikawa M, Yokota H, Fukui et al. Identification of insulin-producing cells derived from embryonic stem cells by zinc-chelating dithizone. Stem Cells 2002;20:284-92.[PubMed]
32. Lukowiak B, Vandewalle B, Riachy R, et al. Identification and purification of functional human beta-cells by a new specific zinc-fluorescent probe. J Histochem Cytochem 2001;49:519-28.[PubMed]
33. Chausmer AB. Zinc, insulin and diabetes. J Am Coll Nutr 1998;17:109-15.[PubMed]
34. Dodson G, Steiner D. The role of assembly in insulin’s biosynthesis. Curr Opin Struct Biol 1998;8:189-94.[PubMed]
35. Sun Y, Chen L, Hou XG, et al. Differentiation of bone marrow-derived mesenchymal stem cells from diabetic patients into insulin-producing cells in vitro. Chin Med J 2007;120:771-6.[PubMed]
36. Kroon E, Martinson LA, Kadoya K, et al. Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat Biotechnol 2008;26:443-52.[PubMed]
[TOP]