Scott L. Pratt, T. Ashley Burns, Erin Curry, Susan K. Duckett
Animal and Veterinary Science Department, Clemson University, Clemson, SC, USA
Correspondence: Scott L. Pratt, Room 117, Poole Agriculture Center, Animal and Veterinary Science Department, Clemson University, Clemson, SC, 29634, USA. E-mail: scottp@clemson.edu
Key words: adipocyte, adipogenesis, microRNA, gene expression, Bos taurus.
Contributions: SLP is the PI and was integral to the total design and execution of the experiments as was Dr. Duckett. In addition, the procedures for the isolation and generation of the stromal-vascular cell cultures were performed by Dr. Pratt. TAB graduate Student of Dr. Duckett’s, adapted published differentiation protocols for bovine primary cultures and suggested using lipid accumulation to verify cell differentiation procedures. EC all qRT-PCR, sub-cloning and sequencing procedures for miRNA were developed by Ms. Curry and utilized on this project, conducted the bioinformatic analysis. Ms. Curry is Dr. Pratt’s graduate student. SKD collaborates on this project, suggested the assessment of miRNA expression in vivo and had tissues of biological interest to analyze, aided in the real time relative expression analysis.
Conflict of interest: the authors report no conflicts of interest.
Received for publication: 28 May 2010.
Revision received: 11 October 2010.
Accepted for publication: 14 October 2010.
©Copyright S.L. Pratt et al., 2010
Licensee PAGEPress, Italy
Journal of Nucleic Acids Investigation 2010; 1:e12
doi:10.4081/jnai.2010.e12
AbstractStudies have recently indicated that the adipogenic process and the expression of genes involved in lipid metabolism may be regulated in part at the post-transcriptional level by a class of small RNA called microRNA (miRNA). The objectives of this study were to i) determine if miRNAs are differentially expressed, and ii) evaluate expression of known miRNA targets in bovine adipocytes. Bovine adipose samples were collected from castrated males fattened on a high concentrate diet (C) or pasture (PA) and were frozen in liquid nitrogen and stored at -80°C, or used to generate primary stromal-vascular cells (SV). SV cells were cultured to confluence (Control) or differentiated at confluence and harvested 2 (D2), 6 (D6), or 12 (D12) days post-confluence. A 3x3 microarray analysis was performed comparing Control and differentiated samples. miR-21, -221, and -222 (P<0.05) were differentially expressed. qRT-PCR was conducted using the in vitro samples, and all three miRNAs were down regulated on D2 (P<0.05). miR-221 and -222 were decreased on D6 compared to Control (P<0.05), but only miR-222 expression was decreased at D12 (P<0.05) compared to Control. miR-21 increased in expression compared to Control on D12 (P<0.05). In vivo, only miR-21 expression was affected and it was reduced in PA compared to C fat samples (P<0.05). Two targets of miR-21 are Programmed Cell Death Protein 4 (PDCD4) and Phosphatase and Tensin Homolog (PTEN), and neither messenger RNA was differentially expressed in vitro (P>0.05), but both messenger RNAs were elevated for PA compared to C (P<0.05). These data show that miRNAs are differentially expressed in adipose cells and tissue, and that miR-21 may be involved in adipocyte function by regulating the translation of PDCD4 and PTEN. |
The majority of research examining changes in lipogenic gene expression has been conducted in rodents, in part because of the availability of tractable in vitro models for adipocyte differentiation and hypertrophy. Most information describing adipogenesis utilizes the 3T3-L1 cell line;1 however, use of primary bovine preadipocyte or clonal preadipocyte cultures has been reported.2,3,4,5 In addition to dietary, hormonal, and/or transcription factor impacts on adipogenesis, microRNAs (miRNAs) are implicated in adipocyte differentiation6,7 and/or adipocyte function.8,9,10
miRNAs are small non-coding RNA that are present and expressed in the cells of all plants and animals examined to date, and regulate gene expression post-transcriptionally by either translational repression or RNA interference (as reviewed by (11)). The understanding of miRNA action became more complicated when investigators established that in quiescent cells, miRNA may actually stimulate protein translation.12,13 miRNA tend to be highly conserved across species,14,15,16,17 and it is predicted that 60% of human protein-coding genes are under selective pressure to conserve miRNA target sites.18 miRNA expression in human and rodent adipocytes, including 3T3-L1 cells6,9 and in bovine adipose tissue19 has been reported; however, no information on the expression differences or role of specific miRNA has been reported for bovine adipogenesis. We hypothesize that miRNA profiles change during adipogenesis for Bos taurus cells, and that adipogenic genes are regulated by the miRNA. In this study we will i) evaluate the identity of miRNA present in bovine adipocytes and their differential expression both in vivo and in vitro, and ii) determine the expression of two specific target messenger RNA of miR-21.
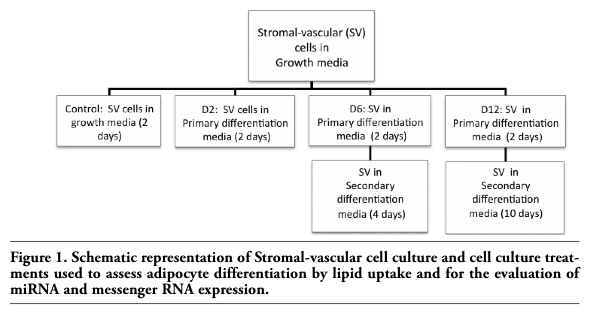
Subcutaneous bovine adipose tissues were harvested at sacrifice from Angus-crossbred castrated males fed a high concentrate diet (C; n=7) or on pasture only (PA; n=6). Samples were harvested from the tail head of the animal, rinsed in PBS, frozen in liquid N2, and stored at -80°C. The C treatment was formulated to contain 94.11% rolled corn, 2.91% soybean meal, 1.50% limestone, 0.95% urea, and 0.53% of trace minerals (dry matter basis; for more information on diet see Pavan et al., 2007).20 Two stromal-vascular cell cultures were established by harvesting flank adipose from a second set of two 18 mo old Angus-crossbred castrated males fattened on pasture and primary cultures established as described by Hirai et al., 2007.3 Tissue was minced under sterile conditions prior to digest with collagenase I in Hanks balanced salt at 37°C with shaking for ~2 h. In addition, cells were expanded in T75 flask, subjected to trypsinization, and frozen to be used at a later date. Growth media (DMEM containing 10% FCS, and 2X antibiotic/antimycotic) and the primary differentiation media (DMEM containing 5% FCS, 2X antibiotic/antimycotic, insulin (2.5 μg/mL), 0.25 μM Dex, 5 μM troglitasone, and 0.5 mM IBMX) were as reported by Hirai et al., 2007.3 Secondary differentiation media (DMEM, 5% FCS, 2X antibiotic/antimycotic, insulin (2.5 μg/mL), and troglitasone (5 μM)) was added to cells after 48 h of culture in primary differentiation media. Culture was maintained in secondary differentiation media for 10 days. To differentiate SV cells, cells were held at confluence for 2 d in growth media. Growth media was removed and primary differentiation media added. The experimental design for cell culture and cell differentiation is shown in Figure 1. All treatments were performed on both primary cultures in duplicate.
To verify that the primary cultures differentiated into adipocytes, lipid analysis was conducted on control stromal-vascular cells and differentiated cells. Cellular fatty acids were extracted and transmethylated according to Folch et al. (1957)21 and Park and Goins (1994),22 respectively. In brief, fatty acids were extracted using 2:1 chloroform:methanol and 0.58% sodium chloride followed by vortexing and centrifugation. The lower layer was then evaporated under nitrogen and subsequently transmethylated with 500 μL hexane and
500 μL hexane containing internal standard (1 μg/μL C23:0). Samples were run on a gas chromatograph, Supelco SP-2560 capillary column, with a split ratio of 5:1 for long chain fatty acids. Percentages, as well as gravimetric weights, were calculated for the predominant fatty acids found in cells and tissues.
Total RNA was isolated from SV control cells (Control) or from differentiated cells at Day 2, 6, or 12 of treatment using the mirVana miRNA Isolation Kit (Ambion, Austin, TX, USA) per manufacturer's protocol. Concentration was determined using the nanodrop spectrophotometer, and quality assessed by the 260/280 ratio and slab gel denaturing electrophoresis.23,24 Total RNA isolation directly from adipose tissue was as described by Duckett, et al., 2009.23
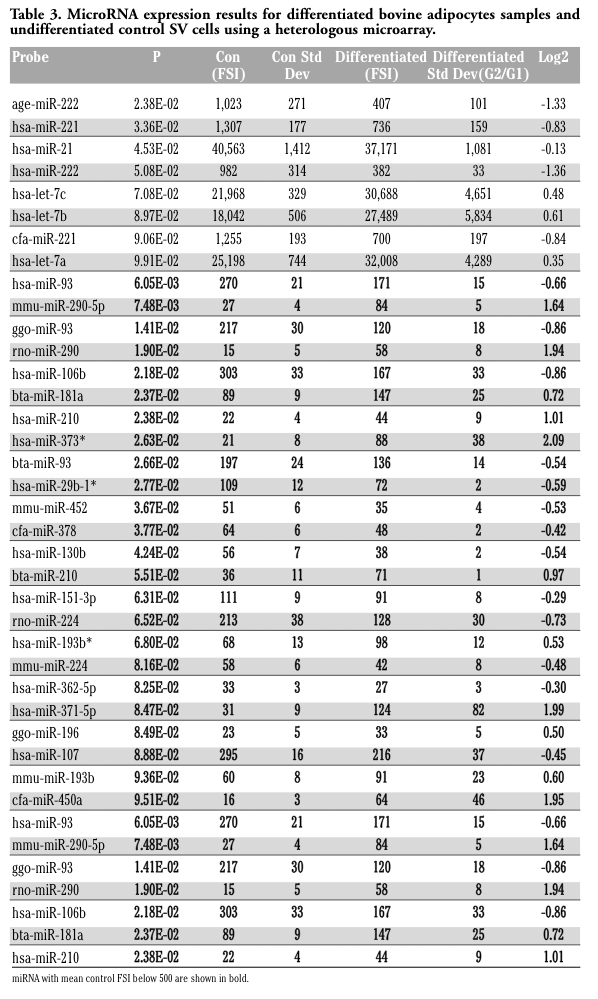
Microarray analysis was performed on heterologous miRNA chips14 using a service provider (LC Sciences, LLC, Houston, TX, USA; www.lcsciences.com) to identify the miRNA profile present in bovine preadipocytes and adipocytes. Control (n=3) and differentiated samples (n=2 from D2; n=1 D6) were labeled with Cy3 and/or Cy5 dyes as described below. Three technical replicates were conducted, each using dual hybridization of a Control and differentiated sample. Briefly, five µg of total cellular RNA was size fractionated using a YM-100 Microcon centrifugal filter (Millipore, Bedford, MA, USA) to isolate small RNAs. Small RNAs less than 300 nt were 3’- extended with a poly(A) tail using poly(A) polymerase. An oligonucleotide tag was ligated to the poly(A) tail for later fluorescent dye staining. Hybridization was performed overnight on a µParaflo microfluidic chip using a micro-circulation pump (Atactic Technologies, Houston, TX, USA). On the microfluidic chip, each detection probe consisted of a chemically modified nucleotide coding segment complementary to a known miRNA target or control RNA and a spacer segment of polyethylene glycol to extend the coding segment away from the substrate. miRNA probes (n=1636) were complementary to known miRNAs from 22 different species, including 125 known bovine miRNA. The detection probes were made by in situ synthesis using photogenerated reagent chemistry. The hybridization melting temperatures were balanced by chemical modifications of the detection probes. Hybridization used 100 µL 6x SSPE buffer (0.90 M NaCl, 60 mM Na2HPO4, 6 mM EDTA, pH 6.8) containing 25.0% formamide at 34°C.
Tag detection was determined using fluorescence labeling with tag-specific dyes. Images were collected using the GenePix® 4000B laser scanner (Molecular Device, Inc, Sunnyvale, CA, USA) and digitized using Array-Pro image analysis software (Media Cybernetics, Silver Spring, MD, USA). Data were analyzed by first subtracting the background and then normalizing the signals using a LOWESS filter (Locally-weighted Regression) to compensate for the intensity difference between Cy3 and Cy5. The ratio of the two sets of detected signals (log2 transformed, balanced) and P-values of the T-test were calculated. Differentially detected signals were those with less than 0.05 P-values.
The signal intensities of the duplicate reactions were averaged and, although the microarray service provider recommended including all samples with detectable signal greater than 30 in analysis, a more conservative threshold was adopted. Detectable transcripts included those with average fluorescent signal intensity (FSI) greater than or equal to 100 and only miRNA with an FSI greater than 500 and predicted or tending to be differentially expressed were chosen for further analysis.
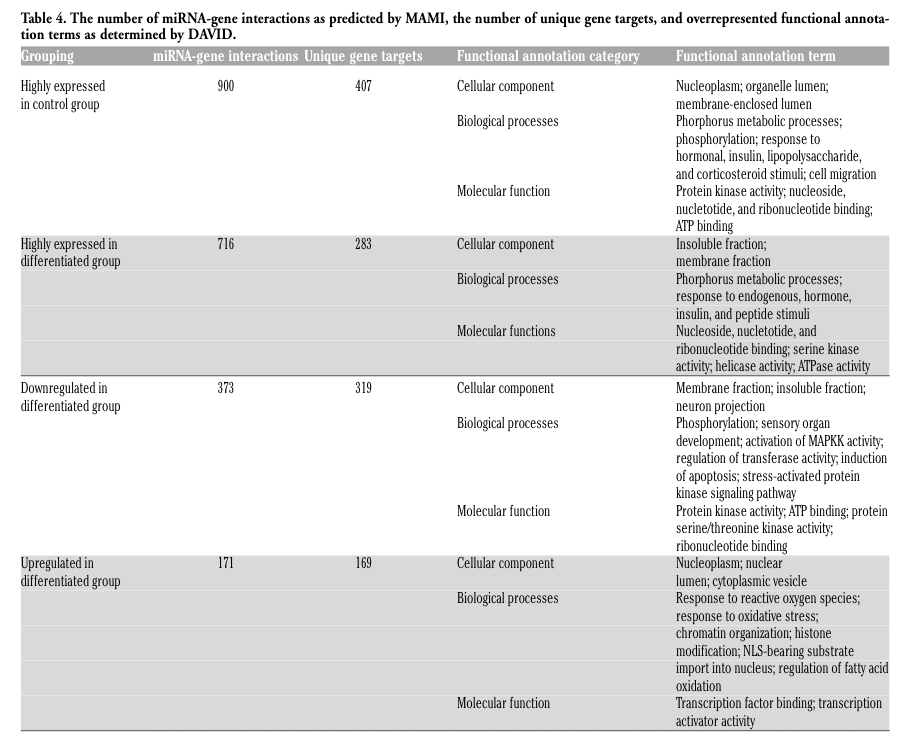
In order to identify the functions of differentially expressed miRNAs between Control and Differentiated groups, the target genes were extracted using MAMI server and database (meta mir: target inference; http://mami.med.harvard.edu/) which provides a composite prediction of target genes from five independent target prediction tools: TargetScan,25 miRanda,26 DIANA-microT,27 miRtarget28 and picTar.29 Targets were predicted for highly expressed miRNAs (FSI>10,000) from Control and Differentiated groups and also those miRNAs differentially expressed between groups (P<0.05). A MAMI sensitivity of 0.66 and a specificity of 0.8 were used to minimize Type I Errors.
The Database for Annotation, Visualization and Integrated Discovery (DAVID; http://david.abcc.ncifcrf.gov;30,31 is a free online bioinformatics resource that provides interpretation of biological themes associated with large gene lists. DAVID was used to annotate MAMI-predicted target genes of differentially expressed miRNAs and to identify significant functional enrichment in the miRNA gene targets relative to the whole genome background. Entrez gene IDs were uploaded to the functional annotation tool and enriched biological themes were analyzed using a modified Fisher's exact test (EASE score; P<0.05), medium classification stringency, minimum enrichment score of 1.3, and a minimum fold enrichment of 1.5. Functional annotation charts and clustering were analyzed to determine enriched molecular functions, biological processes, cellular components, biochemical pathways, and other gene annotations.
End point RT-PCR for the detection of specific miRNAs was conducted on total RNA using the mirVana qRT-PCR Kit (Ambion, Austin, TX, USA) and products subcloned and sequenced.14 All real time qRT-PCR for miRNAs were performed using the same kit and commercially available primers (Ambion). Standard curves were generated for each primer set to determine primer efficiencies. 5S was used as the house keeping gene to normalize miRNA expression data32 and GAPDH was used to normalize messenger RNA data.23 Expression for normalized miRNA and messenger RNA was compared to Control for in vitro all differentiated samples; and PA compared to C for in vivo samples using REST-MCS-b 9 August 2006 release and REST 2009, respectively.33 The REST-MCS-b 9 August 2006 release allows for the comparison of multiple treatments to controls which were performed in the in vitro cell culture experiments. The RNA amounts for each sequence were calculated at the CT at which each fluorescent signal was first detected above background. Normalized CT values for messenger RNA and 5S for miRNA) were calculated for each sample and subjected to ANOVA, as described below. The fold changes in gene expression for PA versus C or Day versus Control for in vivo and in vitro samples, respectively, were calculated by using the X-ΔΔCT method, where X is the primer efficiency for each gene of interest, according to Pfaffl et al. (2002).33 These results are shown in the Figure 2. In addition, normalized qRT-PCR for each messenger RNA and miRNA evaluated, data were subjected to further analysis where cell line, day and cell line*day were main effects using GLM procedures. No cell line*day effect was observed for any miRNA (P>0.05) therefore data for each cell line were pooled. All expression data are presented as fold-difference with standard error as reported by the REST programs from Control or C expression for in vitro and in vivo samples, respectively. RT-PCR procedures for bovine p27, PDCD4 and PTEN was conducted using the same analysis procedures described by Duckett et al., 2009.23 Primer sequences used for detection of messenger RNA are given in Table 1.
Frozen adipose and liver tissue was homogenized in RIPA buffer and protein concentration determined using BioRad reagents (BioRad, Hercules, CA, USA). Cells were harvested, washed in PBS, and extracts generated by suspending cell pellets in RIPA buffer. Bovine liver and adipose; and cell extracts (~50 μg) extracts were subjected to denaturing SDS-PAGE under reducing conditions on 4-15% gradient gels (BioRad). Proteins were transferred on to nitrocellulose, blocked in 5% W/V non-fat dry milk in tris-buffered saline containing 0.1% tween 20, and incubated with primary Ab for β-Actin, PTEN, or PDCD4 (Cell Signaling, Danvers, MA) overnight at 4°C. The β-Actin Ab use chosen to serve as a positive control to detect a protein that should be expressed in liver and adipose tissue. Membranes were washed, incubated with HRP-linked secondary Ab and detection conducted using chemiluminescent reagents and visualized on the Alpha Innotech FC2 system (Santa Clara, CA, USA).
To determine effectiveness of the differentiation treatment of SV cells as outlined in Figure 1, culture was conducted in duplicate on one of the primary SV cultures. Cells were harvested and used to generate samples for lipid content analysis.20,23 Lipid accumulation/ synthesis results are given in Table 2. Control and differentiated D2 samples did not differ in total fatty acid composition or in the percent increase in lipid accumulation, but total fatty acid composition significantly increased by 75.37% and 268.17% in differentiated D6, and D12 treatments (P<0.004 and P<.0.0001, respectively).
After establishing differentiation procedures, we performed microarray analysis on samples to detect miRNA expression differences early in adipogenesis prior to or corresponding to lipid synthesis/accumulation. Depending on label and chip, 156 to 166 miRNAs were detected out of the total 1636 present on the chips. Of the 125 bovine sequences on the chip, 17 to 19 exhibited FSI above the cut off. T-Test analysis showed that a total of 32 miRNAs tended to be or were differentially regulated (Table 3); however, many exhibited a high degree of sequence similarity between species and therefore the real number of unique miRNAs detected and possibly differentially expressed was 27. Only two of the 32 sequences were previously identified Bos taurus miRNA. Further, only sequences for miR-21, -221, and -222 were differentially expressed (P<0.05) and exhibited a hybridization signal above 500 FSI. In addition several miR-21, -221, and -222 sequences (dependant on species) tended to be differentially expressed and further analysis focused on these 3 miRNA. Neither of the two bovine miRNA probes on the chip exhibited an FSI above 500 and no further analysis was conducted. End point RT-PCR analysis, subcloning and sequencing verified the presence of bovine miR-21, -221, and -222 (Figure 3). Furthermore, the predicted messenger RNA targets PTEN and PDCD4 (regulated by miR-21)34,35,36,37,38 and p27 (regulated by miR-221 and -222)39,40,41,42 were detected using end point RT-PCR and products verified by sequencing (Figure 3). The messenger RNAs were present in SV cells, differentiated cells, and in adipose tissue. Other miRNAs detected by array hybridization that were not differentially expressed, but have been previously reported to be involved in adipocyte function and/or adipogenesis were miR-29a, b, and c, miR-143, miR-145, miR-196.6,9,19 miRNA that were not detected as in previous reports were miR-9 and the miR-17-92 cluster.6,10 let-7a, b, and c tended (P<0.10) to be differentially expressed and exhibited a hybridization signal at the level of our conservative cut off of 500 FSI, but were not evaluated further.
Further analysis of the 3x3 array data was conducted extracting predicted targets (based on the human genome) for highly expressed in the Control and Differentiated groups and the miRNA that were down or up regulated in each group using the procedures described in the Design and Methods section. Functional annotation charts and clustering were analyzed to determine enriched molecular functions, biological processes, cellular components, biochemical pathways, and other gene annotations (Table 4). Only the miR-21, 221, and -222 were down regulated, we focused on the biological processes identified in Table 4. These included kinase activity and regulation of apoptosis. Further, a search of published literature established functional targets for all the miRNA chosen for further analysis. For more information view supplementary data.
We next determined if the miRNA or mRNA profiles differed due to i) the differentiation process or ii) due to diet in mature adipocytes completed. For the latter, cell culture experiments were deemed unsuitable due to issues with maintaining a differentiated primary cultures for extended periods of time, therefore, in vivo experiments were conducted on tissues obtained from adipose of castrated males feed C or PA diets. qRT-PCR was conducted for miR-21, -221, -222 and for PTEN and PDCD4 using each verified primer set on in vitro and in vivo samples. Further analysis of p27 was not performed due to the abundance of literature describing p27 function in adipogenesis,44,45,46 and evaluating expression of p27 in mature adipose in vivo would be mute. The expression profile of miR-21, -221, and -222; and PTEN and PDCD4 are shown for in vitro cell culture samples and in vivo bovine adipose tissue expression in Figure 2. In vitro miR-21 expression is initially repressed by -2.35-fold in D2 versus Control (P<0.05), no difference at D6, but increased by 2.07-fold at D12 compared to Control (P<0.05). In contrast, both miR-221 and -222 are down regulated compared to Control at D2 (P<0.05; 3.7- and 5.5-fold, respectively), and D6 (P<0.05; 1.9- and 3.5-fold, respectively). miR-222 continued to be down regulated compared to Control at D12 (P<0.05). In vivo miR-21 expression was minutely repressed in PA versus C diets (0.2-fold; P<0.05), but neither miR-221 nor -222 expression was altered (P>0.05). Two known messenger RNA targets of miR-21 expression were evaluated at the nucleic acid level. Neither PDCD4 nor PTEN were different form Controls at any time point in vitro (P>0.05); however, both PDCD4 and PTEN were elevated by 3.1- and 4.1-fold respectively in PA versus C samples (P<0.05). Neither PDCD4 nor PTEN were different form Controls at any time point in vitro (P>0.05); however, both PDCD4 and PTEN were elevated by 3.1- and 4.1-fold respectively in PA versus C samples (P<0.05).
Expression data was further verified by performing western blotting of bovine SV cell extracts and adipose tissue extracts. Antibodies generated to β-Actin, PTEN, and PDCD4 (Cell Signaling Technologies Inc., Danover, MA, USA) detected peptides of the expected relative molecular weights when performing SDS-PAGE and immunodetection (Figure 4). The β-Actin antibody detected the ~45 kDa full length peptide and the expected proteolytic clip in bovine liver, and adipose extracts. Antibodies generated to PTEN and PDCD4 detected proteins of 54 and 60 kDa, respectively, in bovine liver and adipose; and in undifferentiated in vitro Control and in pooled differentiated samples.
|
Figure 1. Schematic representation of Stromal-vascular cell culture and cell culture treatments used to assess adipocyte differentiation by lipid uptake and for the evaluation of miRNA and messenger RNA expression. |
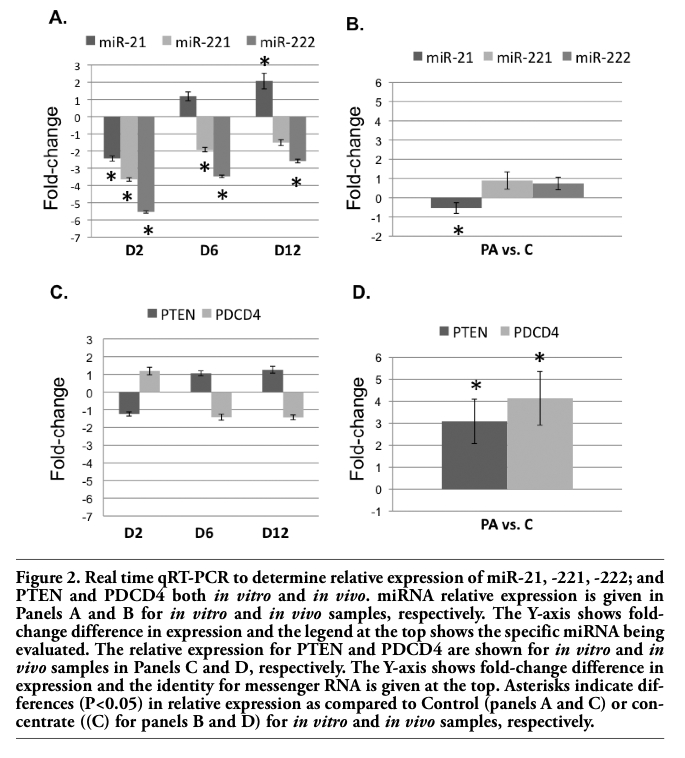 |
Figure 2. Real time qRT-PCR to determine relative expression of miR-21, -221, -222; and PTEN and PDCD4 both in vitro and in vivo. miRNA relative expression is given in Panels A and B for in vitro and in vivo samples, respectively. The Y-axis shows fold-change difference in expression and the legend at the top shows the specific miRNA being evaluated. The relative expression for PTEN and PDCD4 are shown for in vitro and in vivo samples in Panels C and D, respectively. The Y-axis shows fold-change difference in expression and the identity for messenger RNA is given at the top. Asterisks indicate differences (P<0.05) in relative expression as compared to Control (panels A and C) or concentrate ((C) for panels B and D) for in vitro and in vivo samples, respectively. |
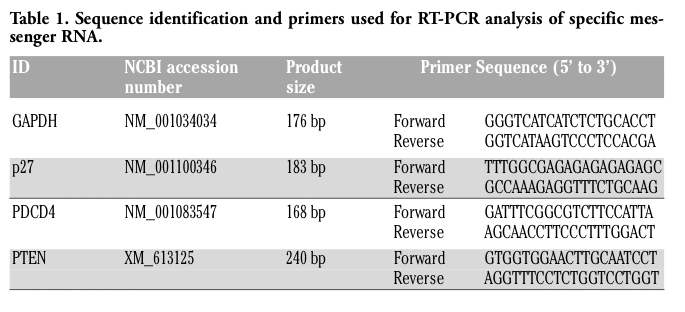 |
Table 1. Sequence identification and primers used for RT-PCR analysis of specific messenger RNA. |
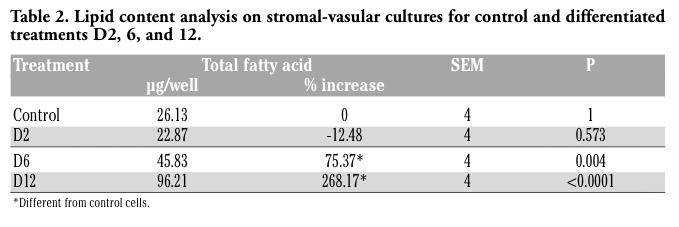 |
Table 2. Lipid content analysis on stromal-vasular cultures for control and differentiated treatments D2, 6, and 12. |
|
|
Literature implicates miRNA involvement in adipogenesis.6,7,47 Hackl et al., 20058 using computational analysis of the 3’ UTR of messenger RNA encoding proteins known to be involved in lipid metabolism predicted that ~71% could possibly be regulated post-transcriptionally by miRNA. Further evidence as to the importance of miRNA in gene regulation is that the molecules tend to be highly conserved across species, and it is predicted that miRNA target sequences are under selective pressure to retain sequence identity for regulation.18 Therefore, miRNA tend to have conserved function and targets across species, which is particularly important when considering that a single miRNA can interact with multiple messenger RNA target sequences and that a specific messenger RNA could be regulated by multiple miRNA (Table 4). Currently, little is known about specific targets for most miRNA as a single miRNA species could target multiple messenger RNA species. Our goal was to evaluate miRNA expression during adipogenesis and in mature adipocyte function. An in vitro primary cell culture system was utilized to study the adipogenic process. However, due to the required extended culture times for mature adipocyte generation, we decided to utilize bovine adipose tissue when evaluating the possible alteration of miRNA expression in mature adipocytes. To achieve the stated objectives we i) isolated and cultured bovine preadipocytes, and ii) verified that the bovine preadipocytes differentiated to adipocytes. We were successful in isolating two bovine SV cultures that were morphologically identical to preadipocytes reported in the literature.2,3,48,49 Further, these cells differentiated into adipocytes as determined by morphology (data not shown), and by lipid uptake (Table 2). Significant lipid accumulation was not observed until D6 of differentiation and we decided to use D2 and D6 total RNA samples to compare to Control samples for array analysis to identify miRNAs that may be differentially expressed in the earlier phases of adipogenesis. Using the verified in vitro adipogenesis model, we identified the presence of multiple miRNA using a heterologous microarray. Table 3 gave the identities of 32 miRNA detected and additional analysis using the MAMI server and database and DAVID, revealed that differentially expressed miRNA (down regulated) were involved in phosphorylation events and apoptosis. The three miRNA that were differentially expressed were miR-21, -221, -222 (P<0.05). miR-221 and -222 are involved in cell cycle regulation via the phosphoprotein p27.39,40,49,50 miR-21 is involved in cell hypertrophy and apoptosis.34,36,51,52,53 Our data show these miRNA are repressed during early bovine adipogenesis, in vitro. miR-21, miR-221 and miR-222 expression was confirmed both by end-point (Figure 2) and qRT-PCR (Figure 3). When confirming and expanding the expression analysis using qRT-PCR, miR-21 increased when comparing D12 to Control which would theoretically decrease the expression miR-21 messenger RNA targets present in differentiating adipocytes and SV. Further, the qRT-PCR expression results were in complete agreement with the microarray data showing that miR-21, -221, and -222 were down regulated early during adipogenesis. However, the degree of differential expression was estimated to be greater using qRT-PCR. Since we did not have an in vitro system that was capable of evaluating mature adipocyte function for extended periods of time, we evaluated the expression of miR-21, -221, and -222 in mature adipoctyes in vivo by assessing their expression levels in bovine adipose tissue of animals fattened on two separate diets.23,54 In comparing diets, miR-221 and -222 showed no difference in their expression in mature adipocytes and this may not be any surprise as these tissues were harvested from 18 month old animals and after nine months of age in Bos taurus, the majority of adipogenesis is due to hypertrophic growth of adipocytes, not adipogenesis.55,56 Interestingly, miR-21 is actually decreased in PA versus C treatment (P<0.05) and miR-21 expression has been implicated in other tissues as important for hypertrophic growth.51 Our in vitro and in vivo data would indicate that the higher levels of miR-21 present in C and D12 samples, repectively would allow for hypertrophic growth required for the increased storage of lipid in adipose tissue54 and differentiated SV cells (Table 2).
We then identified expression of miR-21, 221, and -222 messenger RNA targets of interest present in the in vitro and in vivo models. A search of the literature clearly shows that miR-221 and -222 regulate p2739,40,42 and this protein has been shown to impact early adipogenesis via cell cycle regulation.44,45,46 In fact, both miR-221 and -222 impact the expression of the cyclin-dependant kinase inhibitors p27 and p57.39,40,41,42,57 miR-221 has been shown to be expressed in adipocytes of other species,6,9 but miR-222 has not. However, experiments using in vitro cell models other than adipocytes clearly show a role for miR-221 and -222 in cell cycle regulation.39,40,41,42 These data were generated in cancerous cell lines, but direct effects of miR-221 and -222 on the production of p27 and/or p57 were shown. The role of p27 during adipogenesis is to block cell cycle progression prior to clonal expansion, at which point it is degraded by calpain.45 These data would be consistent with our observations, as levels of miR-221 and 222 decreased in D2 samples, the production of p27 would hypothetically increase, and cells would be inhibited from cell cycle progression.
We decided to focus more on the role of miR-21 in adipogenesis, as it was observed to be the more abundantly expressed (Table 3) and the aspect of apoptosis and/or hypertrophic function in adipocyte development is lacking in the literature. A search of the literature did show that miR-21 impacts two peptides involved in apoptosis, being PTEN and PDCD4.35,36,37,58,59 We then verified the expression of PTEN and PDCD4 in both the in vitro and in vivo systems at the messenger RNA and protein levels. PTEN and PDCD4 are expressed at the nucleic acid (Figure 2) and the protein levels (Figure 4) for both in vitro cell culture and the in vivo mature adipocyte cells. qRT-PCR analysis showed that neither PDCD4 or PTEN expression is altered during the differentiation process of SV to adipocyte, but miR-21 could still alter the protein levels as it increases through adipogenesis. Further research is required to test this possibility. In contrast, diet greatly impacted the expression of PTEN and PDCD4 in mature adipocytes in vivo (Figure 3). In contrast to miR-21, the PTEN and PDCD4, known apoptotic proteins,35,58 are highest in PA samples. This could allow for the scenario where PTEN and PDCD4 could be regulated through diet and a miRNA known to regulate both is also regulated inversely by diet. The result would be that in high concentrate diets, PTEN and PDCD4 are decreased at the messenger RNA level and miR-21 is increased allowing for cell survival by blocking cell death protein expression and allowing for hypertrophic growth required for increased lipid accumulation. This would be similar to findings in other cell/specie systems showing miR-21 blocking the anti-tumor apoptotic effect of PDCD4.34,36,37,38,52,60 To our knowledge this is the first report establishing the expression of PTEN and PDCD4, messenger RNAs encoding two proteins associated with carcinogenesis and cell death,34,35,58,61,62,63 being present in adipose tissue of any species. In addition to the aforementioned hypothesis, PTEN has already been implicated in insulin signaling. PTEN regulates insulin sensitivity through its modulation of the PI3 pathway both in vivo and in vitro.64,65,66,67 The experiments reported here do not expand on the role of PTEN in PI3 signaling nor how it would relate to our hypothesis concerning a role in hypertrophic growth. These data could also indicate that miR-21 is a marker for mature adipocytes as it is present in adipose tissue and its expression increases in vitro as SV cells differentiate into functional adipocytes beginning approximately day 6 of culture.
In conclusion, our data clearly indicate that miRNAs are expressed in bovine SV cells, adipocytes, and adipose tissue as are specific miRNA targets known to be involved in cell cycle, and cell survival. These data also demonstrate that miRNAs are differentially expressed during the adipogenic process, but at present, the regulators of the miRNA expression discussed here are unknown. Further, all miRNAs indentified as being differentially regulated could have impacts on adipogenesis and/or lipid metabolism, but further work to identify specific effects is required.
1. Ntambi JM, Young-Cheul K. Adipocyte differentiation and gene expression. J Nutr 2000;130:3122S-6S.[PubMed]
2. Grant AC, Ortiz-Colon G, Doumit ME, Buskirk DD. Optimization of in vitro conditions for bovine subcutaneous and intramuscular preadipocyte differentiation. J Anim Sci 2008;86:73-82.[PubMed]
3. Hirai S, Matsumoto H, Hino N, et al. Myostatin inhibits differentiation of bovine preadipocyte. Domest Anim Endocrinol 2007;32:1-14.[PubMed]
4. Taniguchi M, Guan LL, Zhang B, et al. Gene expression patterns of bovine perimuscular preadipocytes during adipogenesis. Biochem Biophys Res Commun 2008;366:346-51.[PubMed]
5. Taniguchi M, Guan LL, Zhang B, et al. Adipogenesis of bovine perimuscular preadipocytes. Biochem Biophys Res Commun 2008;366:54-9.[PubMed]
6. Esau C, Kang X, Peralta E, et al. MicroRNA-143 regulates adipocyte differentiation. J Biol Chem 2004;279:52361-5.[PubMed]
7. Wang Q, Li YC, Wang J, et al. miR-17-92 cluster accelerates adipocyte differentiation by negatively regulating tumor-suppressor Rb2/p130. Proc Natl Acad Sci U S A 2008;105:2889-94.[PubMed]
8. Hackl H, Burkard TR, Sturn A, et al. Molecular processes during fat cell development revealed by gene expression profiling and functional annotation. Genome Biol 2005;6:R108.[PubMed]
9. Kajimoto K, Naraba H, Iwai N. MicroRNA and 3T3-L1 pre-adipocyte differentiation. RNA 2006;12:1626-32.[PubMed]
10. He A, Zhu L, Gupta N. Overexpression of micro ribonucleic acid 29, highly up-regulated in diabetic rats, leads to insulin resistance in 3T3-L1 adipocytes. Mol Endocrinol 2007;21:2785-94.[PubMed]
11. Cuellar TL, McManus MT. MicroRNAs and endocrine biology. J Endocrinol 2005;187: 327-32.[PubMed]
12. Vasudevan S, Tong Y, Steitz JA. Switching from repression to activation: microRNAs can up-regulate translation. Science 2007;318:1931-4.[Abstract]
13. Vasudevan S, Tong Y, Steitz JA. Cell-cycle control of microRNA-mediated translation regulation. Cell Cycle 2008;7:1545-9.[PubMed]
14. Curry E, Ellis SE, Pratt SL. Detection of porcine sperm microRNAs using a heterologous microRNA microarray and reverse transcriptase polymerase chain reaction. Mol Reprod Dev 2009;76:218-9.[PubMed]
15. Sawera M, Gorodkin J, Cirera S, Fredholm M. Mapping and expression studies of the mir17-92 cluster on pig chromosome 11. Mamm Genome 2005;16:594-8.[PubMed]
16. Bentwich I, Avniel A, Karov Y, et al. Identification of hundreds of conserved and nonconserved human microRNAs. Nat Genet 2005;37:766-70.[Abstract]
17. Coutinho LL, Matukumalli LK, Sonstegard TS, et al. Discovery and profiling of bovine microRNAs from immune-related and embryonic tissues. Physiol Genomics 2007;29:35-43.[PubMed]
18. Friedman RC, Farh KK, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009;19:92-105.[PubMed]
19. Gu Z, Eleswarapu S, Jiang H. Identification and characterization of microRNAs from the bovine adipose tissue and mammary gland. FEBS Lett 2007;581:981-8.[PubMed]
20. Pavan E, Duckett SK. Corn oil supplementation to steers grazing endophyte-free tall fescue. II. Effects on longissimus muscle and subcutaneous adipose fatty acid composition and stearoyl-CoA desaturase activity and expression. J Anim Sci 2007;85:1731-40.[PubMed]
21. Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem 1957;226:497-509.[PubMed]
22. Park P, Goins RE. In situ preparation of fatty acid methyl esters for analysis of fatty acid composition in food. J Food Sci 1994;59:1262-6.[Abstract]
23. Duckett SK, Pratt SL, Pavan E. Corn oil or corn grain supplementation to steers grazing endophyte-free tall fescue. II. Effects on subcutaneous fatty acid content and lipogenic gene expression. J Anim Sci 2009;87:1120-8.[PubMed]
24. Pratt SL, Anthony RV. The growth hormone receptor messenger ribonucleic acid present in ovine fetal liver is a variant form. Endocrinology. 1995;136:2150-5.[PubMed]
25. Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005;120:15-20.[PubMed]
26. John B, Enright AJ, Aravin A, et al. Human MicroRNA targets. PLoS Biol 2004;2:e363.[PubMed]
27. Kiriakidou M, Nelson PT, Kouranov A, et al. A combined computational-experimental approach predicts human microRNA targets. Genes Dev 2004;18:1165-78.[PubMed]
28. Grun D, Wang YL, Langenberger D, et al. microRNA target predictions across seven Drosophila species and comparison to mammalian targets. PLoS Comput Biol 2005;1:e13.[PubMed]
29. Krek A, Grun D, Poy MN, et al. Combinatorial microRNA target predictions. Nat Genet 2005;37:495-500.[PubMed]
30. Dennis G, Jr., Sherman BT, Hosack DA, et al. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol 2003;4:P3.[PubMed]
31. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 2009;4:44-57.[PubMed]
32. Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol Lett 2004;26:509-15.[PubMed]
33. Pfaffl MW, Horgan GW, Dempfle L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 2002; 30:e36.[PubMed]
34. Asangani IA, Rasheed SA, Nikolova DA, et al. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008;27:2128-36.[PubMed]
35. Frankel LB, Christoffersen NR, Jacobsen A, et al. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J Biol Chem 2008;283:1026-33.[PubMed]
36. Lu Z, Liu M, Stribinskis V, Klinge CM, et al. MicroRNA-21 promotes cell transformation by targeting the programmed cell death 4 gene. Oncogene 2008;27:4373-9.[Abstract]
37. Meng F, Henson R, Wehbe-Janek H, et al. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007;133:647-58.[PubMed]
38. Zhu S, Wu H, Wu F, et al. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res 2008;18:350-9.[PubMed]
39. Fornari F, Gramantieri L, Ferracin M, et al. MiR-221 controls CDKN1C/p57 and CDKN1B/p27 expression in human hepatocellular carcinoma. Oncogene 2008;27:5651-61.[PubMed]
40. Galardi S, Mercatelli N, Giorda E, et al. miR-221 and miR-222 expression affects the proliferation potential of human prostate carcinoma cell lines by targeting p27Kip1. J Biol Chem 2007;282:23716-24.[PubMed]
41. le Sage C, Nagel R, Egan DA, et al. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J 2007; 26:3699-708.[PubMed]
42. Visone R, Russo L, Pallante P, et al. MicroRNAs (miR)-221 and miR-222, both overexpressed in human thyroid papillary carcinomas, regulate p27Kip1 protein levels and cell cycle. Endocr Relat Cancer 2007;14:791-8.[PubMed]
43. Pezzolesi MG, Platzer P, Waite KA, Eng C. Differential expression of PTEN-targeting microRNAs miR-19a and miR-21 in Cowden syndrome. Am J Hum Genet 2008; 82:1141-9.[PubMed]
44. Lin J, Della-Fera MA, Li C, et al. P27 knockout mice: reduced myostatin in muscle and altered adipogenesis. Biochem Biophys Res Commun 2003;300: 938-42.[PubMed]
45. Patel YM, Lane MD. Mitotic clonal expansion during preadipocyte differentiation: calpain-mediated turnover of p27. J Biol Chem 2000;275:17653-60.[PubMed]
46. Auld CA, Morrison RF. Evidence for cytosolic p27(Kip1) ubiquitylation and degradation during adipocyte hyperplasia. Obesity (Silver Spring) 2006;14:2136-44.[PubMed]
47. Wang MY, Grayburn P, Chen S, et al. Adipogenic capacity and the susceptibility to type 2 diabetes and metabolic syndrome. Proc Natl Acad Sci USA 2008;105:6139-44.[PubMed]
48. Ohsaki H, Sawa T, Sasazaki S, et al. Stearoyl-CoA desaturase mRNA expression during bovine adipocyte differentiation in primary culture derived from Japanese Black and Holstein cattle. Comp Biochem Physiol A Mol Integr Physiol 2007;148:629-34.[PubMed]
49. Gillies JK, Lorimer IA. Regulation of p27Kip1 by miRNA 221/222 in glioblastoma. Cell Cycle 2007;6:2005-9.[PubMed]
50. le Sage C, Nagel R, Agami R. Diverse ways to control p27Kip1 function: miRNAs come into play. Cell Cycle 2007ì;6:2742-9.[PubMed]
51. Cheng Y, Ji R, Yue J, et al. MicroRNAs are aberrantly expressed in hypertrophic heart: do they play a role in cardiac hypertrophy? Am J Pathol 2007;170:1831-40.[PubMed]
52. Chan JA, Krichevsky AM, Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res 2005;65:6029-33.[PubMed]
53. Corsten MF, Miranda R, Kasmieh R, et al. MicroRNA-21 knockdown disrupts glioma growth in vivo and displays synergistic cytotoxicity with neural precursor cell delivered S-TRAIL in human gliomas. Cancer Res 2007;67:8994-9000.[PubMed]
54. Pavan E, Duckett SK, Andrae JG. Corn oil supplementation to steers grazing endophyte-free tall fescue. I. Effects on in vivo digestibility, performance, and carcass traits. J Anim Sci 2007;85:1330-9.[PubMed]
55. Baldwin RL, Reichl JR, Louis S, et al. Effects of age, pregnancy, and lactation on rat, guinea pig, and cow adipose enzyme activities and cow adipose metabolism. J Dairy Sci 1973;56:340-9.[PubMed]
56. Hood RL, Allen CE. Cellularity of bovine adipose tissue. J Lipid Res 1973;14:605-10.[PubMed]
57. Medina R, Zaidi SK, Liu CG, et al. MicroRNAs 221 and 222 bypass quiescence and compromise cell survival. Cancer Res 2008;68:2773-80.[PubMed]
58. Chen Q, Wang C, Jiang C, Chen D. Exogenous PTEN gene induces apoptosis in breast carcinoma cell line MDA468. J Huazhong Univ Sci Technolog Med Sci 2007;27:61-4.[PubMed]
59. Li X, Lin G, Wu B, et al. Overexpression of PTEN induces cell growth arrest and apoptosis in human breast cancer ZR-75-1 cells. Acta Biochim Biophys Sin (Shanghai) 2007;39:745-50.[PubMed]
60. Zhu S, Si ML, Wu H, Mo YY. MicroRNA-21 targets the tumor suppressor gene tropomyosin 1 (TPM1). J Biol Chem 2007; 282:14328-36.[PubMed]
61. Blanco-Aparicio C, Renner O, Leal JF, Carnero A. PTEN, more than the AKT pathway. Carcinogenesis 2007;28:1379-86.[PubMed]
62. Dreher T, Zentgraf H, Abel U, et al. Reduction of PTEN and p27kip1 expression correlates with tumor grade in prostate cancer. Analysis in radical prostatectomy specimens and needle biopsies. Virchows Arch 2004;444:509-17.[PubMed]
63. Maehama T. PTEN: its deregulation and tumorigenesis. Biol Pharm Bull 2007;30:1 624-7.
64. Nakashima N, Sharma PM, Imamura T, et al. The tumor suppressor PTEN negatively regulates insulin signaling in 3T3-L1 adipocytes. J Biol Chem 2000;275:12889-95.[PubMed]
65. Butler M, McKay RA, Popoff IJ, et al. Specific inhibition of PTEN expression reverses hyperglycemia in diabetic mice. Diabetes 2002;51:1028-34.[PubMed]
66. Jiang G, Zhang BB. Pi 3-kinase and its up- and down-stream modulators as potential targets for the treatment of type II diabetes. Front Biosci 2002;7:d903-7.[PubMed]
67. Kurlawalla-Martinez C, Stiles B, Wang Y, et al. Insulin hypersensitivity and resistance to streptozotocin-induced diabetes in mice lacking PTEN in adipose tissue. Mol Cell Biol 2005;25:2498-510.[PubMed]
[TOP]